
As aves apresentam uma série de adaptações anatômicas acumuladas ao longo de sua história evolutiva. Sacos aéreos e ossos pneumáticos, por exemplo, ajudam a reduzir o peso corporal e a aumentar a eficiência do sistema respiratório; essas características são essenciais para o voo, mas também tornam as aves mais suscetíveis a infecções respiratórias. A evolução das aves, portanto, revela um equilíbrio delicado entre custos e benefícios, permitindo que esses animais vivam no limite de suas capacidades.
Este é o resumo de uma palestra que tive a oportunidade de ministrar no 20º Avistar (Encontro Brasileiro de Observação de Aves), em Maio de 2026. Te convido a conhecer um pouco mais sobre a anatomia das aves e suas especializações para o voo!
Penas
Contradizendo o título deste texto, vou começar falando sobre a anatomia externa das aves e sua característica mais marcante: a presença de penas. As penas são compostas de queratina, a mesma proteína de que são feitas nossas unhas e cabelos. Existe uma grande variedade de tipos de penas, mas somente algumas são utilizadas para voar. Embora as penas sejam indispensáveis para o voo das aves, fósseis indicam que essas estruturas surgiram muito antes do voo, já estando presentes em alguns dinossauros, com funções como proteção, comunicação e camuflagem.
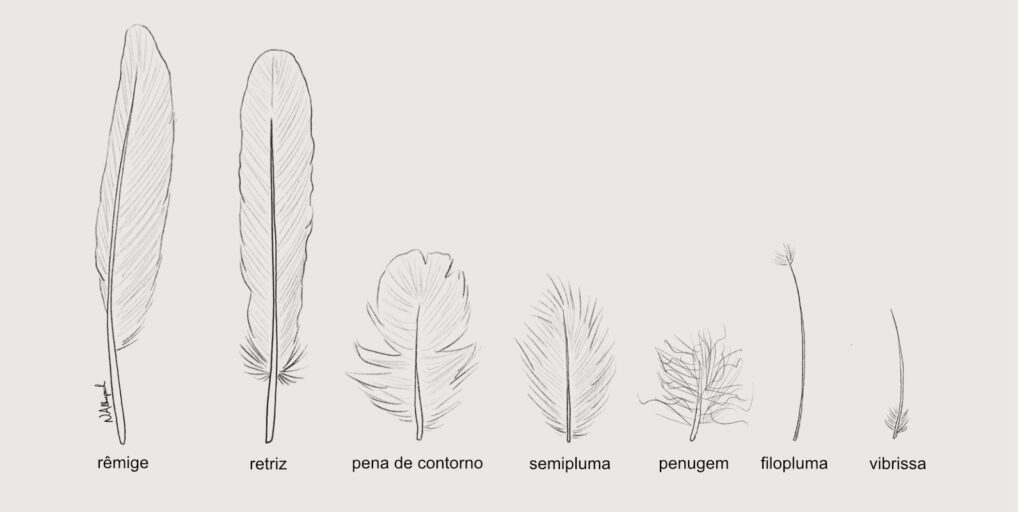
Penas são extremamente leves e resistentes. De maneira semelhante a função dos pelos nos mamíferos, as penas protegem as aves do frio, do sol e de escoriações. Suas cores são importantes na comunicação entre as aves, em especial durante o período reprodutivo, assim como na camuflagem contra predadores. Uma fileira de penas longas e fortes (chamadas de rêmiges) amplia a área de sustentação e confere um perfil chamado “aerofólio” às asas, permitindo o voo. As longas penas da cauda (retrizes) também têm função aerodinâmica.

As penas podem repelir a água. Essa propriedade é relacionada a disposição das barbas e bárbulas das penas de contorno, aliada aos óleos produzidos pela glândula uropigeana, que são cuidadosamente espalhados com o bico sobre toda a plumagem.


Ausência de dentes
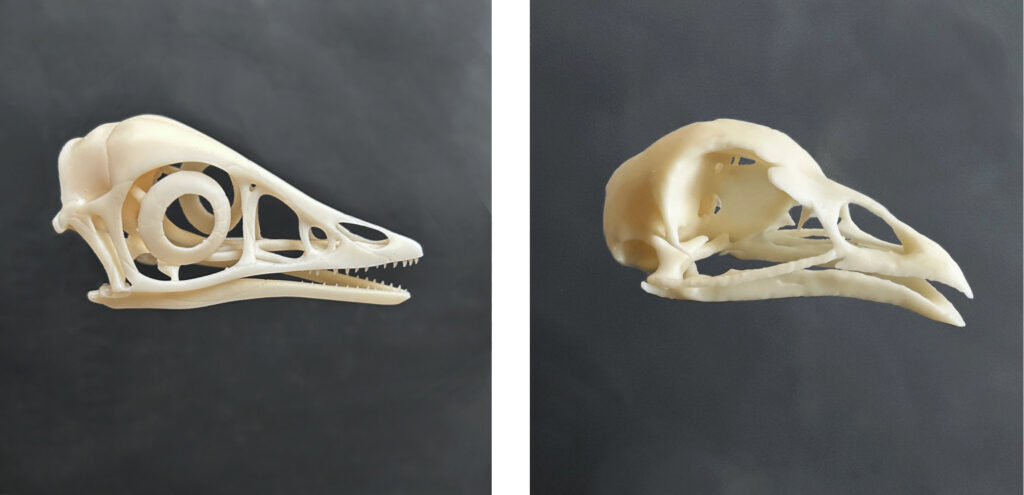
Aves jurássicas, como o famoso Archaeopteryx, apresentavam dentes. Mas nas aves modernas, que vivem nos dias de hoje, não encontramos dentes. Junto com o desenvolvimento de um bico córneo (olha a queratina aqui de novo), acredita-se que a perda dos dentes seja uma das muitas adaptações para o voo, pois representa uma redução no peso corporal da ave.

Um bico que funciona como um radiador → O bico do tucano é extremamente vascularizado e a ave pode controlar o fluxo de sangue nesses vasos. Quando o ambiente está muito quente, mais sangue é enviado para os vasos do bico, que são mais superficiais, facilitando as trocas de calor. Assim o sangue retorna para o resto do corpo ligeiramente mais frio, evitando o superaquecimento da ave. Em dias frios, ou a noite, acontece o contrário: menos sangue é enviado ao bico, evitando que a ave perca calor para o ambiente desnecessariamente.
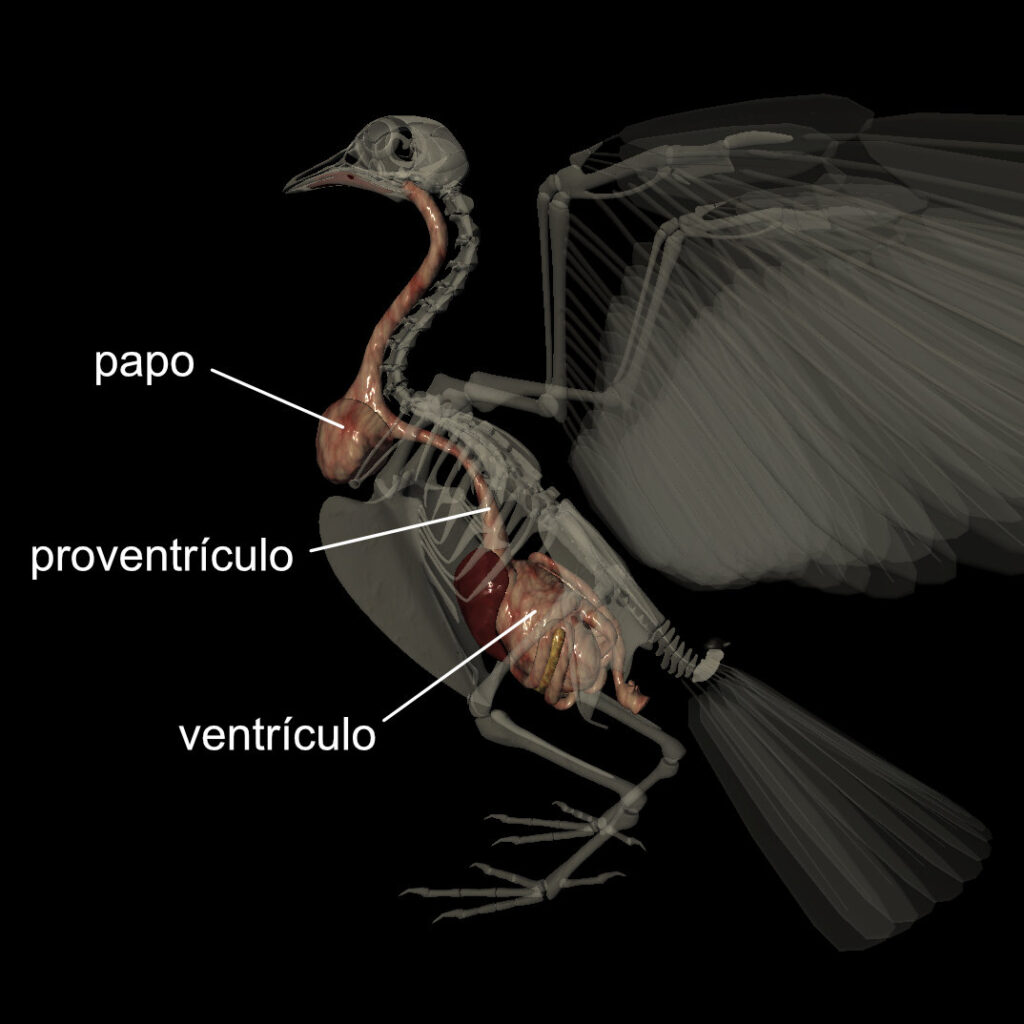
A ausência de dentes, no entanto, trouxe um problema para as aves modernas: como mastigar o alimento? A “solução” surgiu ao longo do tubo digestivo, em um estômago que é dividido em dois órgãos separados: o proventrículo e o ventrículo. O proventrículo funciona de maneira semelhante ao nosso estômago: ele realiza a digestão química, por meio da secreção de enzimas e ácido clorídrico. Já o ventrículo, conhecido popularmente como moela, é responsável pela digestão mecânica. O ventrículo apresenta uma parece muscular bem desenvolvida e seu interior é abrasivo. Essa parede muscular é mais desenvolvida em aves granívoras, cuja dieta exige melhor trituração do alimento para maior aproveitamento dos nutrientes. Para maximizar a digestão mecânica, algumas destas aves ingerem pedrinhas, que ficam alojadas no ventrículo e auxiliam na “mastigação”.
A moela das aves carnívoras separa e compacta os restos não digeridos do alimento (como ossos, pelos, penas e carapaças de insetos) em pelotas que são regurgitadas. Tecnicamente, essas pelotas são chamadas de egagrópilas. Aves que consomem frutos também produzem egagrópilas, formadas por partes mais duras de seu alimento, como cascas e sementes. Ao regurgitar, as aves evitam que o material que não será digerido passe por todo o sistema digestório, livrando-se desse peso adicional logo no início da digestão.


Apesar da ausência de dentes, a digestão nas aves é um processo rápido. As pelotas da foto acima foram expelidas por bem-te-vis aproximadamente 15 minutos após a ingestão dos frutos, durante um experimento feito em cativeiro. Nas aves frugívoras, a passagem do alimento restante por todo o sistema digestório, até a defecação, pode levar menos de uma hora, sendo ainda mais rápida nas aves menores e um pouco mais lenta em aves maiores. Já em aves de rapina, a egagrópila pode demorar algumas horas para ser expelida, geralmente ocorrendo a formação de uma egagrópila por refeição. A defecação também demora um pouco mais nestas aves, em comparação com as aves frugívoras. Em galinhas, que são aves granívoras, o tempo de passagem do alimento pelo sistema digestório leva entre 5 e 6 horas.
Massa corporal centralizada
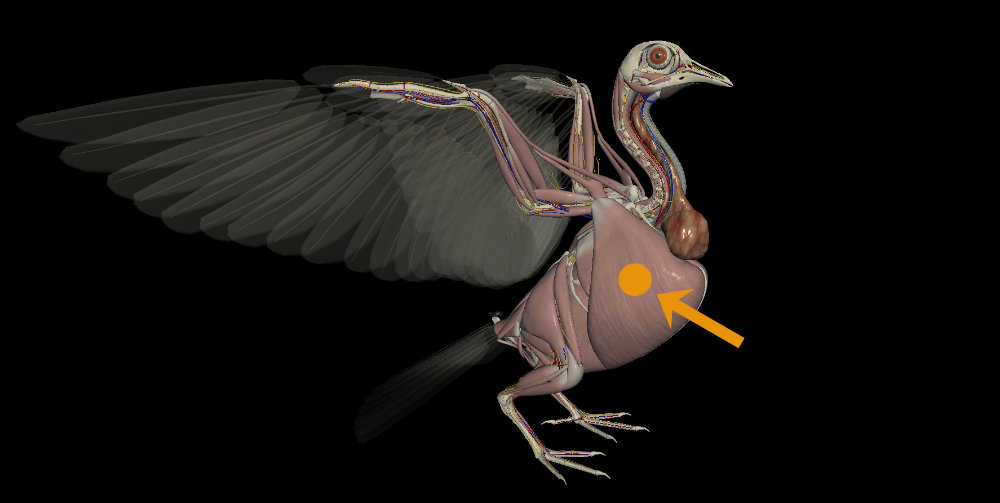
Nas aves, os grandes e potentes músculos do voo estão concentrados na região do peito. Estes músculos estão próximos à linha central do corpo e sua força é transmitida a partir de longos tendões. A musculatura presente nos membros (asas e pernas) é mínima. Desta maneira, o centro de massa corporal (vulgo centro de gravidade) está localizado entre as asas, ligeiramente rebaixado.
A centralização da massa corporal também se deve à redução do peso da cabeça (graças ao bico córneo e ausência de dentes) e da cauda. Os répteis ancestrais das aves apresentavam vértebras na cauda, que foram suprimidas ao longo da evolução e deram origem ao pigóstilo. O pigóstilo é um pequeno osso que resultou da fusão das últimas vértebras caudais, fornecendo uma base resistente para a inserção das penas da cauda (retrizes).
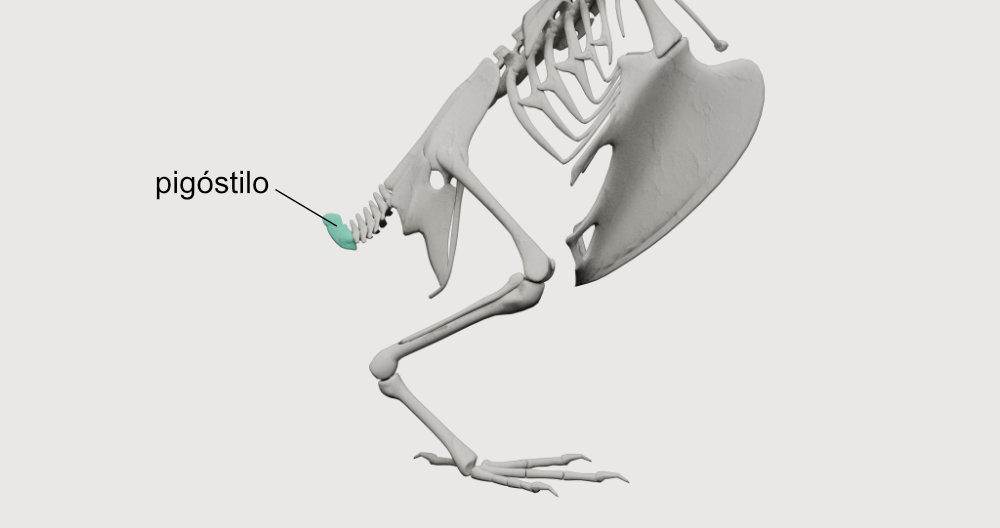
A cauda das aves tem função aerodinâmica: oferece estabilidade e controle para o voo, assim como sustentação durante curvas e voo lento. A cauda também pode ser muito útil em outros contextos, como na seleção sexual (o exemplo do pavão é provavelmente o mais famoso). Em aves escaladoras de troncos, como os pica-paus e os arapaçus, a cauda é de grande importância como apoio, sendo formada por retrizes duras e robustas.

Direita: Benedito-de-testa-amarela (Melanerpes flavifrons) utilizando a cauda como apoio.
Fusão, redução e adaptação dos ossos
O esqueleto das aves é leve, o que permite não tão somente o voo, como também carregar ovos durante sua formação no oviduto das fêmeas, assim como chocar os ovos sem quebrá-los. Isso se dá graças a presença de muitos ossos pneumáticos, que apresentam espaços preenchidos de ar em seu interior. Esses espaços são conectados ao sistema respiratório das aves, tendo ligação com as vias aéreas, os sacos aéreos ou os pulmões. Aves que não voam apresentam ossos com menor grau de pneumatização.
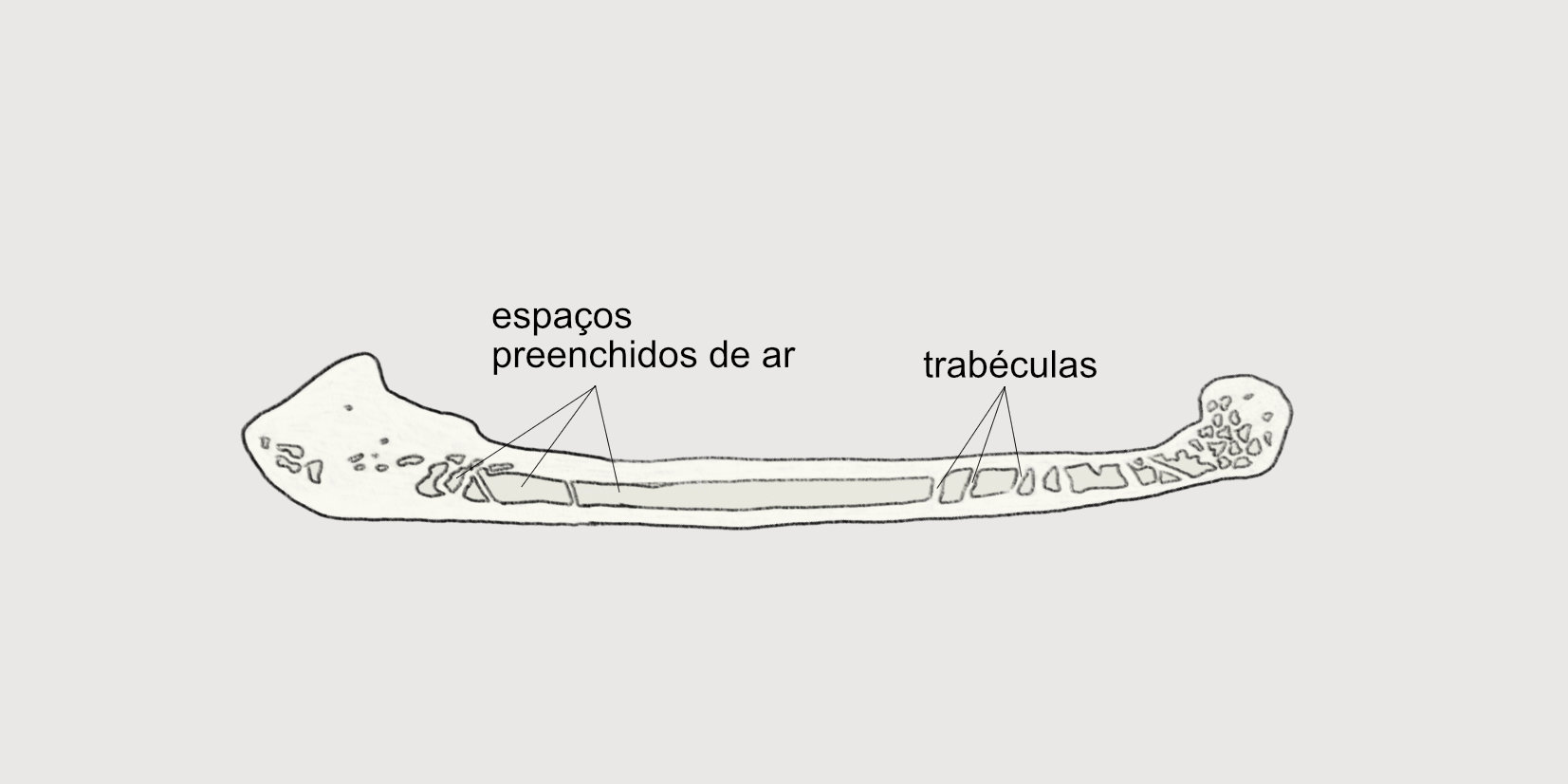
Em comparação com os mamíferos, as aves apresentam um número menor de ossos constituindo seu esqueleto. Por exemplo, humanos adultos têm aproximadamente 206 ossos e cachorros adultos têm cerca de 319 ossos, enquanto uma galinha adulta tem apenas 120 ossos.
Nas aves, muitos ossos foram perdidos ao longo da evolução, ou se fundiram com outros ossos. A fusão de ossos confere mais rigidez para algumas partes do corpo da ave, como tórax e quadril, reduzindo a musculatura que seria necessária para estabilização durante o voo. A fusão de ossos, porém, reduz a flexilidade da ave.
Os ossos do crânio de uma ave adulta também são tão fusionados que constituem praticamente um único osso. Quase não encontramos linhas ou suturas entre eles, como vemos no crânio de um mamífero típico. Essa fusão de ossos craniais confere maior proteção mecânica contra impactos.
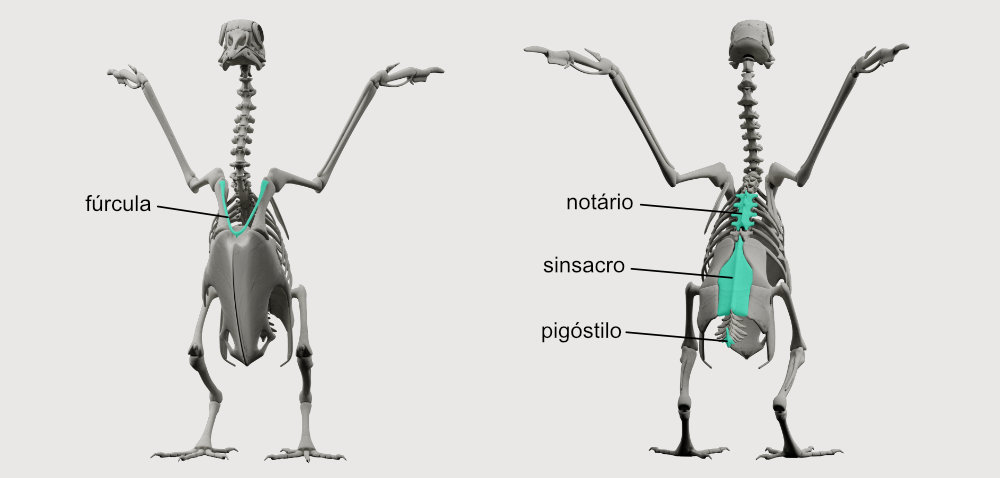
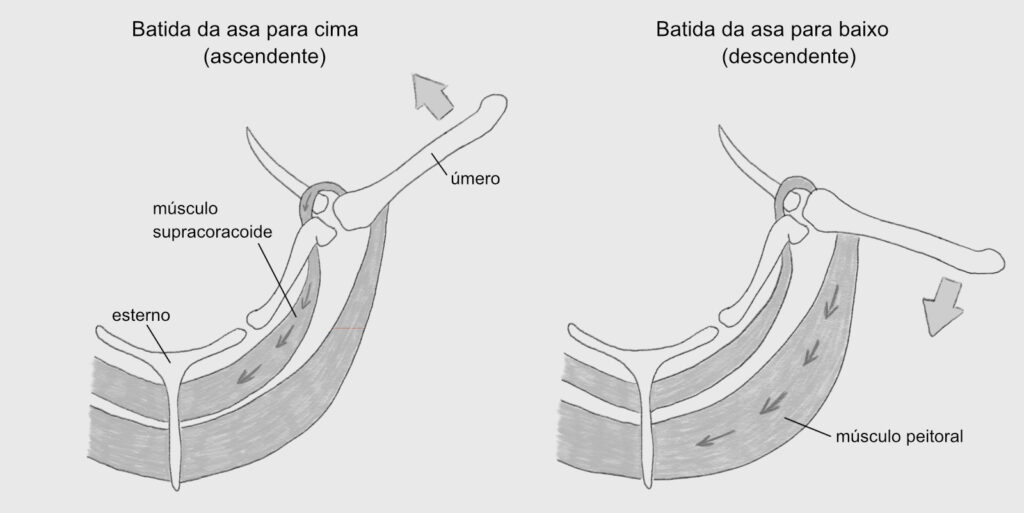
Fúrcula → É o chamado “osso da sorte”, que corresponde a fusão das clavículas. Esse osso age como uma mola, transmitindo forças acumuladas durante o batimento da asa para cima (um movimento que exige menos energia) para o batimento da asa para baixo (um movimento com maior demanda energética).
Notário → Osso resultante da fusão de algumas vértebras torácicas, estabilizando essa parte da coluna vertebral. É encontrado em galinhas, pombas, cegonhas e flamingos, por exemplo.
Sinsacro → Osso que resultou da fusão de vértebras toráxicas, lombares, sacrais e caudais, unido também aos ossos do quadril.
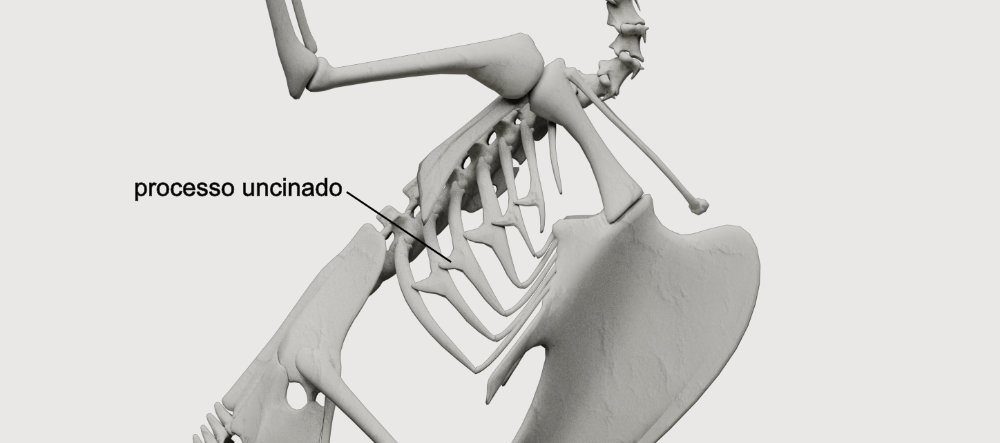
Processos unciados nas costelas → São estruturas em forma de gancho encontradas nas costelas das aves e em alguns répteis. Consistem em pontos adicionais de inserção de músculos, tornando a caixa toráxica mais resistente.
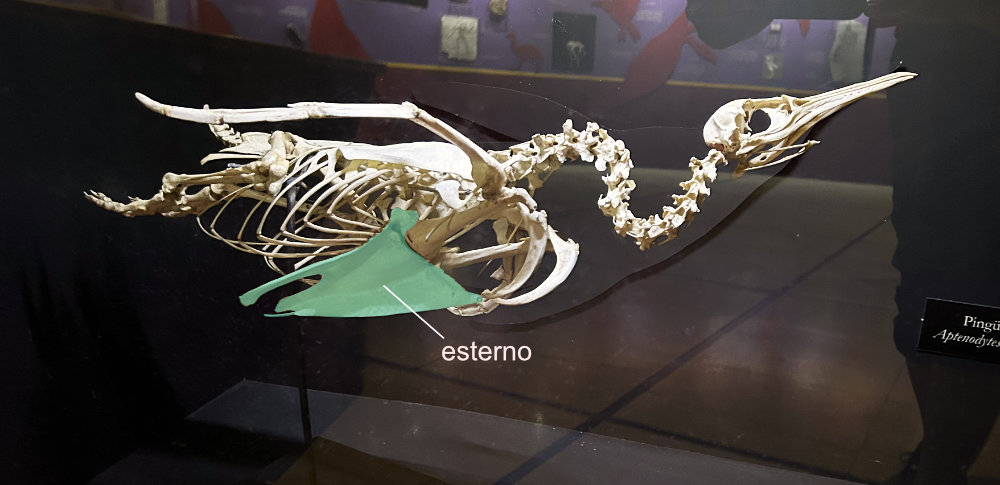
Esterno com quilha → As aves são divididas em carinatas (com quilha, também chamada de carina) e ratitas (sem quilha). A quilha representa uma extensão da área para inserção dos músculos do voo. Quando maior a quilha, maior o tamanho dos músculos peitorais inseridos e maior a capacidade de voo da ave em questão.


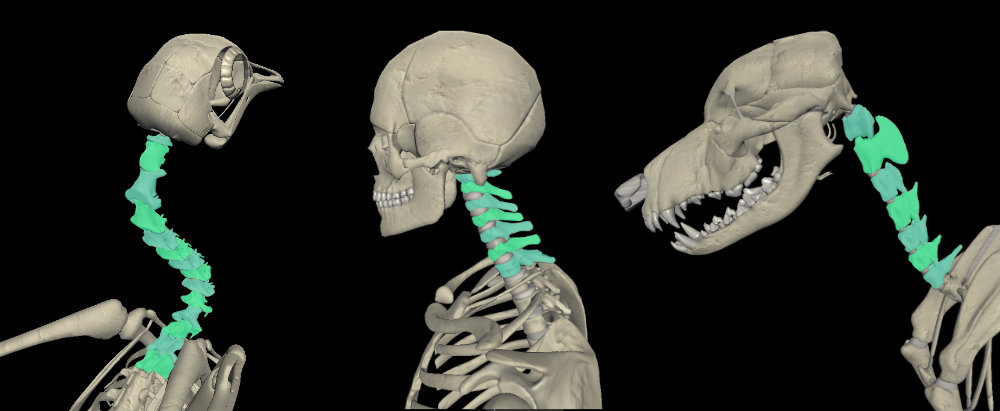
Vértebras cervicais em número variável → Neste caso não houve redução do número de ossos, mas sim um aumento. O longo pescoço das aves compensa a falta de mobilidade das vértebras fusionadas. Passeriformes têm o pescoço mais curto (entre 10 e 15 vértebras), enquanto um cisne pode ter até 25 vértebras cervicais. Um pescoço longo e flexível permite que as aves possam limpar suas penas com o bico e facilita a manipulação dos alimentos.

Asas
Os ossos dos membros anteriores das aves também sofreram muitas fusões. Restaram somente três dedos curtos. A álula (que corresponde ao nosso polegar) é responsável pelo controle fino do voo. Os principais músculos do voo (músculo peitoral e músculo supracoracoide) ligam-se ao úmero.
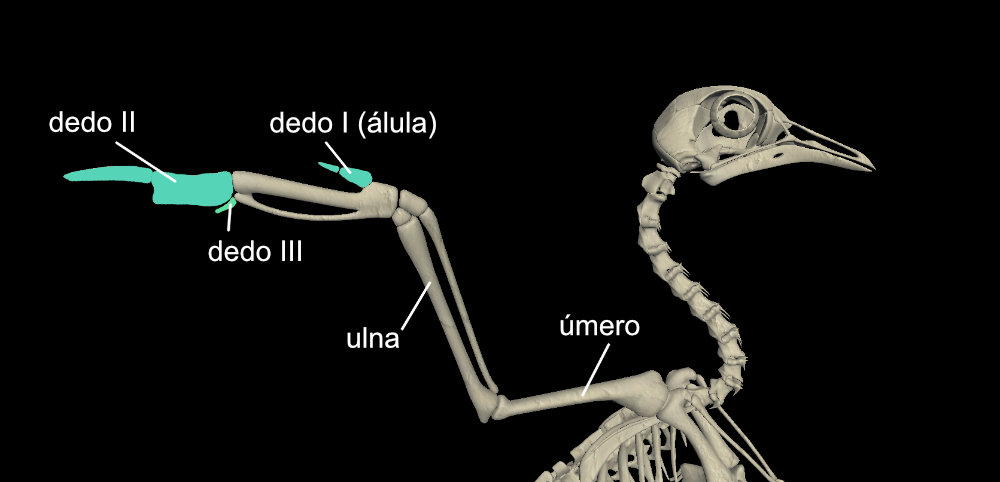
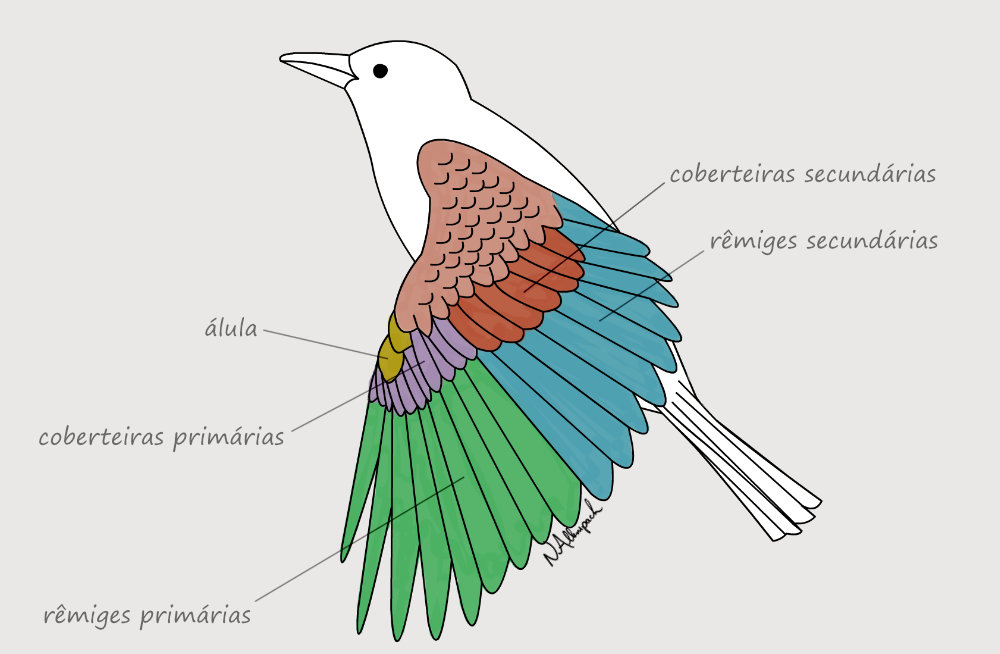
As penas das asas são divididas em dois grupos principais: rêmiges primárias, ligadas aos ossos da “mão”, e rêmiges secundárias, ligadas à ulna. Assim como telhas em um telhado, outros grupos de penas se sobrepõe às rêmiges cobrindo toda a asa. Mas somente as rêmiges são responsáveis por expandir a superfície da asa, tornando o voo possível. As rêmiges são tão essenciais para o voo que precisam ser trocadas gradualmente por novas penas. Assim, a ave pode continuar voando ao longo do processo de muda.
“Desasagem” → Algumas aves aquáticas, como patos, gansos e cisnes, perdem todas as rêmiges de uma vez durante a muda. Desta maneira, essas aves podem ficar até um mês impossibilitadas de voar, período durante o qual ficam mais vulneráveis.
Altas taxas metabólicas
A demanda energética do voo é imensa, assim como a demanda de oxigênio. Por este motivo, o sistema cardiorespiratório das aves precisa ser extremamente eficiente. O coração das aves é proporcionalmente maior que o dos mamíferos, bombeando mais sangue a cada batida. A frequência cardíaca das aves também é maior, quando comparamos aves e mamíferos do mesmo tamanho. O coração de alguns beija-flores pode chegar a mais de mil batimentos por minuto, quando em voo!
Os sacos aéreos são uma característica única das aves, cujos pulmões não se expandem como os nossos. A maior parte das aves têm nove sacos aéreos que ajudam o ar a circular internamente. Eles se ligam aos pulmões, mas também se extendem por dentro de ossos, como o úmero. Mas os sacos aéreos não realizam trocas gasosas, que é função exclusiva do pulmão. No pulmão das aves, o ar flui somente em uma direção. Não há mistura de ar novo e ar já utilizado, como ocorre na maioria dos vertebrados. A quantidade de ar circulando nas aves é mais ou menos constante. Desta maneira, ar fresco e oxigenado está em movimento pelo sistema respiratório das aves tanto na inspiração quanto na expiração, aumentando a eficiência das trocas gasosas.
Devido às características únicas de seu sistema respiratório, as aves podem cantar e respirar ao mesmo tempo!
O diafragma, responsável por expandir a cavidade torácica dos mamíferos durante a inspiração, é ausente nas aves.
Redução do sistema urogenital
Durante a maior parte do ano, o sistema reprodutor das aves permanece reduzido e inativo, ocupando pouco espaço dentro da cavidade abdominal. Os órgãos reprodutivos só vão se expandir durante o período reprodutivo, quando são necessários. Na maioria das aves, o sistema reprodutor feminino apresenta somente um ovário e um oviduto, do lado esquerdo. Algumas aves de rapina constituem exceção e apresentam dois ovários e dois ovidutos.
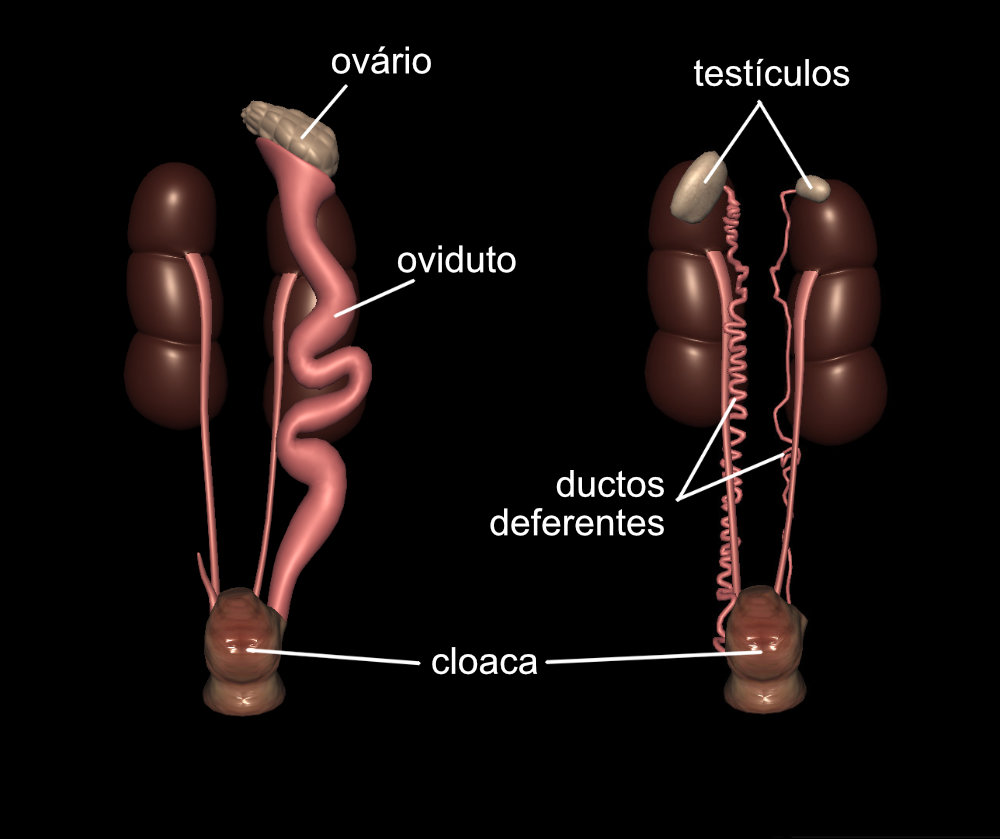
As aves são ovíparas, ou seja, põem ovos. A formação do ovo ao longo do oviduto é rápida, geralmente leva em torno de um dia. Nas galinhas dura entre 24 a 26 horas. A postura geralmente é feita logo pela manhã. Desta maneira, as fêmeas geram um ovo de cada vez, carregando ele dentro de seus corpos pelo menor tempo possível.
Os kiwis são um caso a parte. O ovo, extraordinariamente grande em relação ao tamanho da fêmea, leva cerca de 30 dias para ser formado. Um ovo desse tamanho e que permanecesse tanto tempo dentro do corpo causaria grandes problemas para uma ave típica, mas os kiwis perderam a capacidade de voar e são totalmente terrestres.
A vesícula urinária (vulgo bexiga) é ausente nas aves, com exceção das emas e dos avestruzes. A urina formada pelos rins é levada pelos ureteres até a cloaca. De lá, a urina alcança a parte final do intestino, o cólon e os cecos, onde a água será reabsorvida. O ácido úrico remacescente da urina se mistura com as fezes, que são eliminadas pela cloaca. O ácido úrico corresponde a parte branca das fezes das aves.

A excreção de ácido úrico (dos ditos animais uricotélicos) economiza água e reduz a carga, além de permitir o desenvolvimento embrionário dentro de ovos.
Por fim…
Tentei organizar as informações em diferentes tópicos, mas na prática você deve ter percebido que tudo que apresentei aqui é inter-relacionado. As aves apresentam diversas adaptações anatômicas e fisiológicas que, em conjunto, permitem o voo. Nas espécies que voam longas distâncias, vamos observar essas adaptações em um grau máximo, mas nas espécies que perderam a capacidade de voo podemos vir a encontrar apenas resquícios de algumas dessas adaptações.
Bibliografia consultada
ALLENSPACH, N. A. Frugivoria por aves e fenologia em Miconia albicans e Miconia ligustroides (Melastomataceae), em fragmento de cerrado na região de São Carlos, SP, Brasil. Dissertação (Mestrado em Ecologia e Recursos Naturais) – Universidade Federal de São Carlos, São Carlos, 2009.
BRACHO-ESTÉVANEZ. C. A. et al. Plant traits determine seed retention times in frugivorous birds: Implications for long-distance seed dispersal. Functional Ecology 38, p. 2247-2260, 2024.
FEATHERBASE. Eared Dove – Zenaida auriculata. Disponível em: https://www.featherbase.info/sv/species/zenaida/auriculata. Acesso em fev. 2026.
GALIS, F. Why Do Almost All Mammals Have Seven Cervical Vertebrae? Developmental Constraints, Hox Genes, and Cancer. Journal of Experimental Zoology 285, p. 19-26, 1999.
GASPERIN, G. & PIZO, M. A. Passage time of seeds through the guts of frugivorous birds, a first assessment in Brazil. Revista Brasileira de Ornitologia 20 (1), p. 48‑51, 2012.
GETTY, R. et al. Anatomia dos animais domésticos: vol. 2. Guanabara Koogan. 1986.
GRAYSON, A. D. et al. Determination of Gastrointestinal Transit Times in Barred Owls (Strix varia) by Contrast Fluoroscopy. Journal of Avian Medicine and Surgery 31 (2), p. 123-127, 2017.
KÖNIG, H. E. et al. Avian Anatomy. Textbook and Colour Atlas. Sheffield: 5M Publishing, 2016.
LAWS, J. M. The Laws Guide to Drawing Birds. Berkeley: Heyday, 2012.
LEAL, J. Morfologia aplicada das aves. Uma introdução à Medicina Aviária. Areia: Ed. dos Autores, 2025.
LOVETTE, I. J.; FITZPATRICK, J. W. Handbook of Bird Biology. Ithaca: The Cornell Lab of Ornithology, 2016.
PENDERGRASS, J. How Many Bones Does a Dog Have? A Skeletal System Review. PetLabCo. Disponível em: https://thepetlabco.com/learn/dog/health-wellness/how-many-bones-does-a-dog-have?. Acesso em mar. 2026.
PROCTOR, N. S.; LYNCH, P. J. Manual of Ornithology. Avian Structure & Function. Yale: University Press, 1998.
Schroeder, K. L. et al. Fibre types in primary ‘flight’ muscles of the African Penguin (Spheniscus demersus). Acta Zoologica 96: 510-518, 2015.
SICK, H. Ornitologia Brasileira. Editora Nova Fronteira, 1997.
SVIHUS, B.; ITANI, K. Intestinal Passage and Its Relation to Digestive Processes. Journal of Applied Poultry Research 28 (3), p. 546-555, 2019.
TATTERSALL, G. J. et al. Heat Exchange from the Toucan Bill Reveals a Controllable Vascular Thermal Radiator. Science 325, p. 468-470, 2009. DOI: 10.1126/science.1175553.
TAYLOR, M. The Pocket Book of Bird Anatomy. London: Bloomsbury Wildlife, 2020.
THE CORNELL LAB OF ORNITHOLOGY. All About Feathers. Disponível em: https://academy.allaboutbirds.org/feathers-article/. Acesso mar. 2026.
THE CORNELL LAB OF ORNITHOLOGY. All About Bird Anatomy. Disponível em: https://academy.allaboutbirds.org/features/birdanatomy/. Acesso out. 2025.
THE CORNELL LAB OF ORNITHOLOGY. Birds of the World – Neotropic Cormorant (Nannopterum brasilianum). Disponível em: https://birdsoftheworld.org/bow/species/neocor/cur/introduction?lang=pt_BR. Acesso mai, 2026.
THOMAS, A. L. R. On the Tail of Birds. BioScience 47 (4), p. 215-225, 1997.